https://mp.weixin.qq.com/s/TA-puXA1_Jfha7b5mrg5pA
金鱼属于鲤形目、鲤亚科、鲫属,形态优美,身姿奇异,色彩绚丽,品种繁多,有草金、文、蛋、龙金、龙背等五大类,250个品种之多,是世界著名三大观赏鱼类之一,达尔文形容它有着无限多样的颜色和非凡的表型结构变化。金鱼起源于中国,有“国粹”之称,在中国传统文化中有着和平、美好、吉庆、富有的象征,深受老百姓的喜爱和推崇。早在晋朝(公元265~420年)就有“赤鳞鱼”记载,距今有1700多年的驯养历史,其祖先为野生鲫鱼。在唐代至宋代早期,金鱼被半家化用于行善放生,嘉兴、杭州等地有政府监管行为的放生池,如杭州六和塔寺后山涧池,在浙江多地《地方通志》中均有记载。南宋时期进入家驯化阶段,将金鱼养在堂前屋后的家池中,当时文人以赏析为雅,特别是南宋赵构皇帝对金鱼情有独钟,在杭州德寿宫亲自养殖玩乐。金鱼最早的品种记载是在南宋时期,为白色和花斑。明朝晚期,金鱼逐步养殖到盆或缸中,饲养条件的改变,对金鱼的发育、生理、形态产生了影响,如游行缓慢,产生双尾,背鳍缺失,新品种随之产生。到清朝时期,特别是晚清养殖盛行,进入人工选择阶段,产生龙睛、狮头、鹅头、望天眼、绒球、珠鳞等珍贵品种。世界范围内,金鱼于1502年传到日本,在17世纪末传到英国,18世纪传入欧洲,1874 年传入美国,之后传遍全世界。
图1 福寿(福州)供图
2020年11月2日, PNAS在线发表由福州市多位学者完成的题为“The evolutionary origin and domestication history of goldfish (Carassius auratus)”(金鱼的进化起源和驯养历史)的研究成果,深入探讨了金鱼的起源、演化、驯化和人工选择。福州是传统三大金鱼产区之一,有三百多年的发展历史,金鱼养殖产业发展迅速,有“中国金鱼之都”之称。金鱼的研究对进一步促进福州金鱼产业的发展有着重要的意义。
基于金鱼的重要文化、观赏及科学研究价值,特别是其丰富的变异性,在动植物中相当罕见,课题组从以下几个方面做了深入研究。首先,完善并提升高质量基因组。研究人员使用PacBio单分子测序、染色质三维构象捕获 (Hi-C),结合光学图谱BioNano基因组研究技术,构建了高质量基因图谱,将基因组95.75% 重叠群(contigs)锚定到50条假染色体上,获得单倍型参考基因组,其大小为1.73Gbp,scaffold N50 为 606kb,基因组完整性达94.7%,同时预测了50条染色体中38条的潜在着丝粒区域,更加突出了组装的基因组的完整性。其次,解析金鱼亚基因组及其在鲤亚科中的演化。多倍化现象在脊椎动物中极为罕见,以金鱼祖先鲫为代表的鲤亚科鱼类在演化历史上可能经历了四轮全基因组加倍事件。针对金鱼的基因组是来自于两个祖先物种的杂交事件(也称异源多倍体)还是来自于同源多倍化的问题,科研人员在前人研究基础上,通过重测序6个鲤科代表性二倍体(2n=50)物种:条纹小䰾(Puntius semifasciolatus), 大鳞四须魮(Hypsibarbus vernayi), 长臀鲃(Mystacoleucus marginatus), 黑鳍袋唇鱼(Balantiocheilos melanopterus), 施氏魮(Barbonymus schwanenfdi), 裂峡鲃(Hampala macrolepidota),通过比对率发现,䰾鱼平均80%的读段比对到金鱼的一个染色体簇群上,将金鱼50条染色体明显分开为两个亚组A(ChrA01~A25)、B(ChrB01~B25),且二者基因组大小相近。基于1个核基因的系统进化分析,即结缔组织生长因子基因在二倍体金鱼中仅有1个拷贝,但在四倍体金鱼中含有2个拷贝,系统发育树显示鲃亚科物种的同源基因与金鱼A亚基因组同源基因能聚类到一起(斑马鱼和鲤鱼作外群),验证了之前关于金鱼为四倍体,是异源杂交染色体加倍的假说,而䰾亚科(Barbinae)为最有可能的二倍体祖先种之一,推测金鱼祖先在1000至1200万年前发生过古老的杂交事件。基于同义替换率Ks计算, A、B亚基因组大约分化于14.84-16.67 百万年前,先于金鱼和鲤鱼的分化(11.72-13.16 百万年前),暗示产生异源多倍体金鱼的祖先种早于金鱼与鲤鱼的分化,同时表明全基因组复制(WGD)事件发生在金鱼和普通鲤鱼的共同血统中,这也对他们拥有相同数目染色体的事实的支持。金鱼A、B亚基因组间整体共线性较强,但存在6个同源染色体对较大的片段反转,进一步支持了金鱼是异源四倍体的假设。有意思的是金鱼在Chr4/Chr29上存在高度不共线性的染色体区域,功能富集显示在免疫应答、疾病抗性上起着重要的作用,推测促进了鲤科物种为适应多种病原体而在不同自然条件下栖息和生活。通过金鱼10个不同组织的RNA-seq分析,通过A和B两个亚基因组之间的差异表达,发现同源基因在两个亚基因组之间表现出不对称的表达模式,而全基因组范围内的表达水平总体上更偏好A亚基因组。
图2 金鱼演化示意图
第三,通过群体研究金鱼家训化进程。研究人员对185个具有代表性的金鱼品种和16条野生鲫鱼个体进行重测序,结合33个前人鲤鱼测序数据(三者无生殖隔离),通过SNP位点构建系统发育树显示,金鱼与鲫鱼的亲缘关系高于鲤鱼。主成分分析(PCA)将鲫鱼与金鱼明显分开,进一步佐证了金鱼起源于鲫鱼的假设。组成分分析、系统发育树、群体结构分析、基因流分析等多重证据显示,所有的金鱼品种共享一定数量的血缘,金鱼经历了野生鲫鱼到草金,接着发展到文种,再产生蛋种的家训化历史,而三分法(草系、蛋系和文系)更符合金鱼的进化历史。连锁不平衡及遗传多样性分析也进一步证实上述结论,金鱼驯化过程中种群遗传瓶颈的存在和强烈的人工选择,从鲫鱼和普通金鱼的驯化过程中遗传多样性的增加,表明了从鲫鱼驯化后,金鱼重要遗传变异的积累。此外,研究人员将遗传变异与许多典型特征(眼睛、背鳍、鳞片、颜色)联系起来,通过遗传分化指数(FST)分析,发现50个受选择区域,选择性清除分析发现393个受选择的基因中,有13个基因与斑马鱼中存在敲除系基因同源,这些敲除系基因与行为 (pcdh15a和agrp)、眼部萎缩(ndufs5)、细胞迁移(nav3和zap70)或脑部萎缩(aurka)等相关。推测在整个金鱼群体中这些基因有可能是目标性状在选择过程中的直接靶标,也有可能参与金鱼的主要原始驯化性状有关的表型分化。第四,金鱼重要典型表型性状的组学研究。利用全基因组关联分析(GWAS)定位到与透明鳞相关的基因(Cau.B15G0000010),为酪氨酸蛋白激酶受体基因,这是一个在中国遗传学研究史上著名的孟德尔性状,为不完全显性(T/ t基因座),该基因早在1928年被陈桢教授描述过,该基因在斑马鱼中与色素细胞的发育有关,受体酪氨酸激酶家族的成员在人类黑色素瘤的发展中起作用。其邻近基因编码β-分解酶1(bace1)基因,该基因参与斑马鱼黑素细胞迁移,这可能解释了透明鳞片表型通常与金鱼的黑色素瘤表型相关的原因。背鳍是区分文种和蛋种分类的主要标志,研究人员通过文种和蛋种杂交后代F1背鳍性状分离比,暗示了背鳍特征受多个基因位点控制,并具有母体遗传效应。关联分析鉴定到13条染色体上的8.96 Mb区域上,378个与背鳍表型具有相关性的基因,包括背侧纵向血管(itgb1a,rab13和e2f8),背侧异常(kif5ba),背主动脉异常(fev,cxcr4b,gpr183a,tal1,grlx2和uchl5),腹侧(zic2a)和弯曲背鳍(atp1a2a和dhfr),这些基因可能在从文种金鱼通过人工选择驯化至蛋种金鱼以及背鳍发育中发挥了关键作用。本研究中发布的高质量金鱼基因组和多样性数据,为金鱼作为大量候选功能基因的模型以及广泛的家养物种进化案例研究提供了新的知识和资源。图3 金鱼透明鳞相关性状的全基因组关联分析a,文种和蛋种;b,文种和蛋种的群体主成分分析;c, 正对透明鳞性状的曼哈顿图;d,一般线性模型下GWAS的分位数图;e,位于ChrB15染色体上候选基因位置;f,候选区域的基因型分析;g,候选基因及-UTR调控元件
总的来说,本研究提供了高质量基因组,为金鱼分子育种提供基础,有助于推动品种多样性选择和产业的发展。通过亚基因组的研究,为研究异源多倍体脊椎动物的基因组演化和功能提供了新的思路;通过对金鱼系列表型性状的挖掘与解析,为解析脊椎鱼类多样性的演化机制提供多的研究线索,有助于将金鱼成为脊椎动物生理生化和进化优良遗传模型,来探讨脊椎动物突变的遗传机制。
据悉,该研究由福建师范大学陈由强教授、福建农林大学张积森教授联合发起,福建师范大学生命科学学院、福建农林大学基因组与生物技术研究中心、闽江学院、比利时根特大学等单位协作完成。福建师范大学博士生陈多为第一作者,福建农林大学博士生张清、闽江学院唐唯其博士、福建师范大学黄镇讲师、福建农林大学王刚博士为共同第一作者。福建农林大学的张积森教授为本研究通讯作者,福建师范大学陈由强教授、福建农林大学唐海宝教授,比利时根特大学Van de Peer教授、福建师范大学黄镇讲师为共同通讯作者。



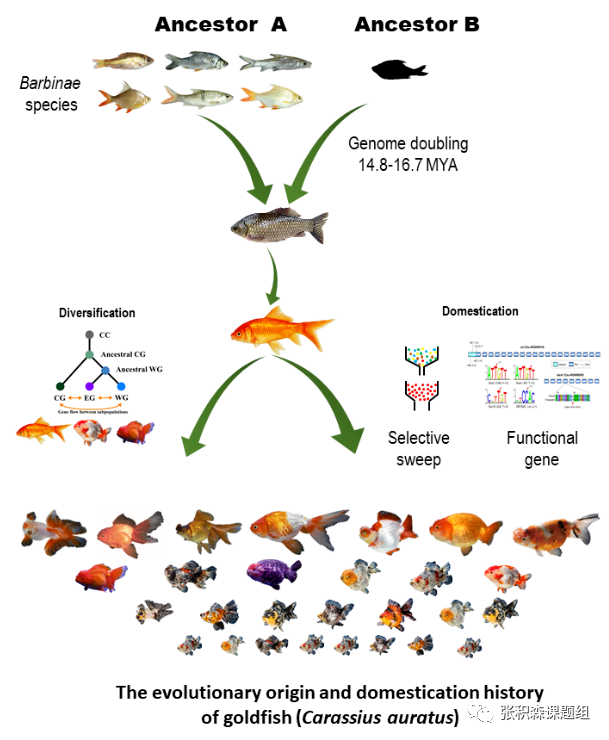
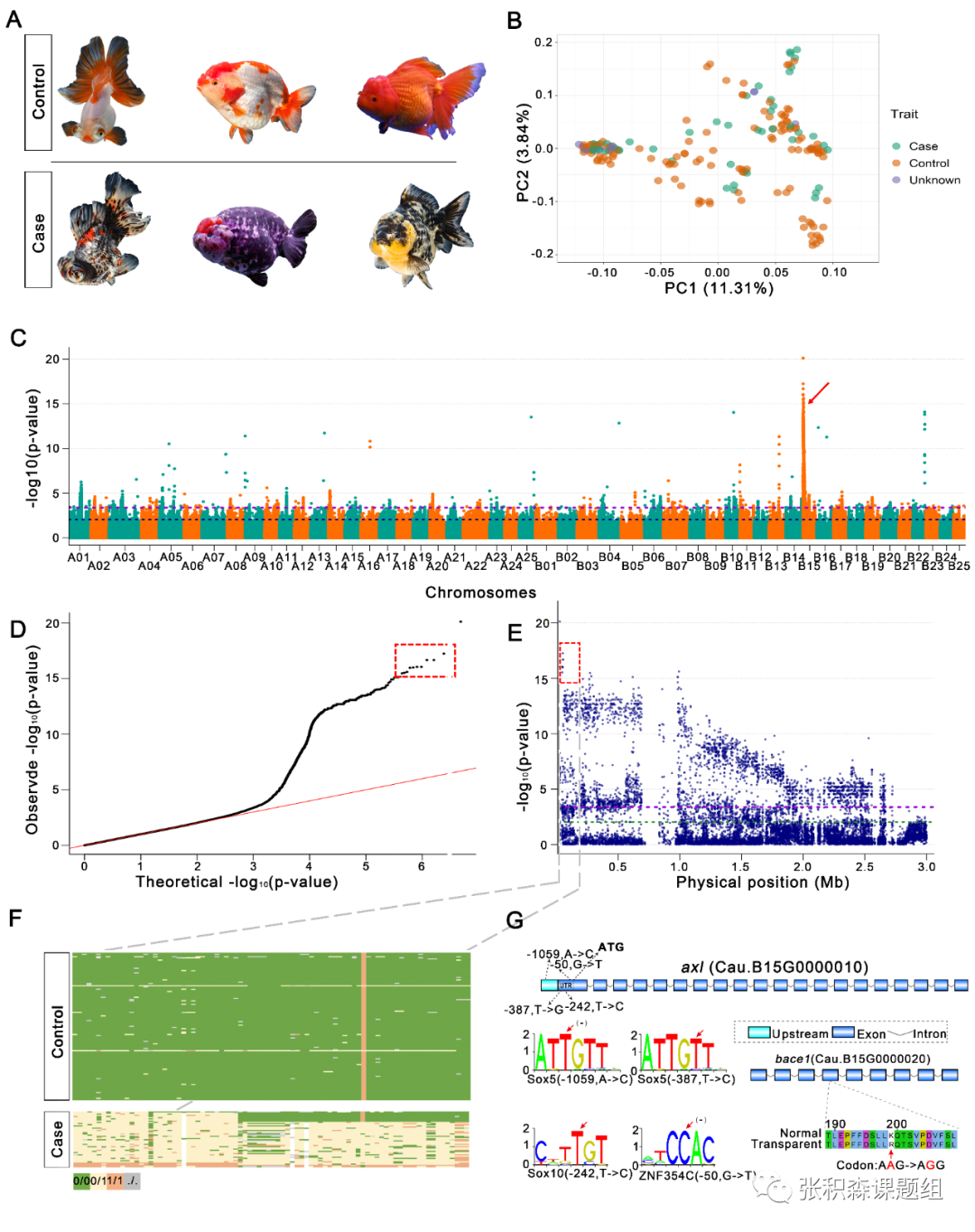 图3 金鱼透明鳞相关性状的全基因组关联分析a,文种和蛋种;b,文种和蛋种的群体主成分分析;c, 正对透明鳞性状的曼哈顿图;d,一般线性模型下GWAS的分位数图;e,位于ChrB15染色体上候选基因位置;f,候选区域的基因型分析;g,候选基因及-UTR调控元件
图3 金鱼透明鳞相关性状的全基因组关联分析a,文种和蛋种;b,文种和蛋种的群体主成分分析;c, 正对透明鳞性状的曼哈顿图;d,一般线性模型下GWAS的分位数图;e,位于ChrB15染色体上候选基因位置;f,候选区域的基因型分析;g,候选基因及-UTR调控元件